
As Richard Feynman points out, every 'why' question in science needs to be treated with caution. This is because there are always several different levels at which a why problem can be answered, depending on what kind of response you're looking for.
Taking Feynman's illustrative example, if pizza arrives at your door and your partner asks why pizza has come, you could fully answer the question in any of the following ways:
- Because I was hungry so I ordered pizza.
- Because someone drove the pizza here from the pizza shop.
- Because in a capitalistic society, you can exchange money for pizza.
- Because I have UberEats on my phone, so I can order food whenever I'm hungry.
And so on and so forth. The same goes for why questions in biology - we can go pretty deep if we try hard enough.
So like any good 5 year old, in this article I'll do just that and keep asking why until we get somewhere interesting.
Table of Contents
- Bee stingers are fishhooks
- A biological poison pump
- Super-organisms, the immune system and colonial life
- Group selection
- Kin selection and biological altruism
- Indirect fitness and the haplodiploidy hypothesis
- The haplodiploidy hypothesis isn't perfect
- Conclusion
Bee stingers are fishhooks
First, we need to be clear about what is actually happening when a honey bee stings you. Rather than a clean hypodermic needle, a honey bee's stinger is actually covered in barbs.
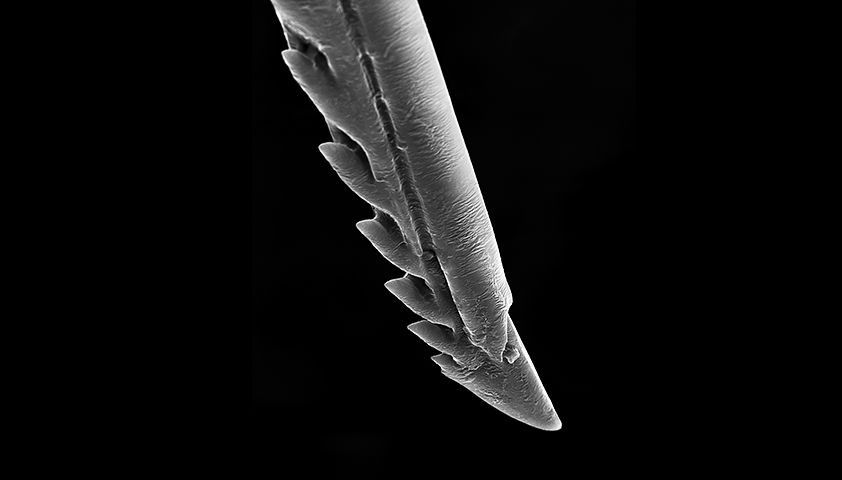
Looks like a very fancy fishhook right? Except it's more gruesome than a fishhook, because after a bee has stung you, it tries to fly away. The barbs keep its stinger in place and the result is the picture at the top of this article. Ouch.
So at a first glance, the answer to the main question is easy. A honey bee dies when it stings you because its stinger is covered in barbs, causing its abdomen to get ripped out when it tries to fly away. And surviving with your guts spilling out everywhere is pretty bloody hard.
But why on Earth would evolution have favoured such a suicidal mechanism? Isn't natural selection supposed to favour survival of the fittest not survival of the suicidal?
A biological poison pump
If we take a look at the entire mechanism of the honey bee stinger we can begin to understand why this suicidal strategy is so effective.
Although the ripping out of the stinger does cause the bee to eventually die, in this process, a venom sack along with a muscular pump are also left behind.
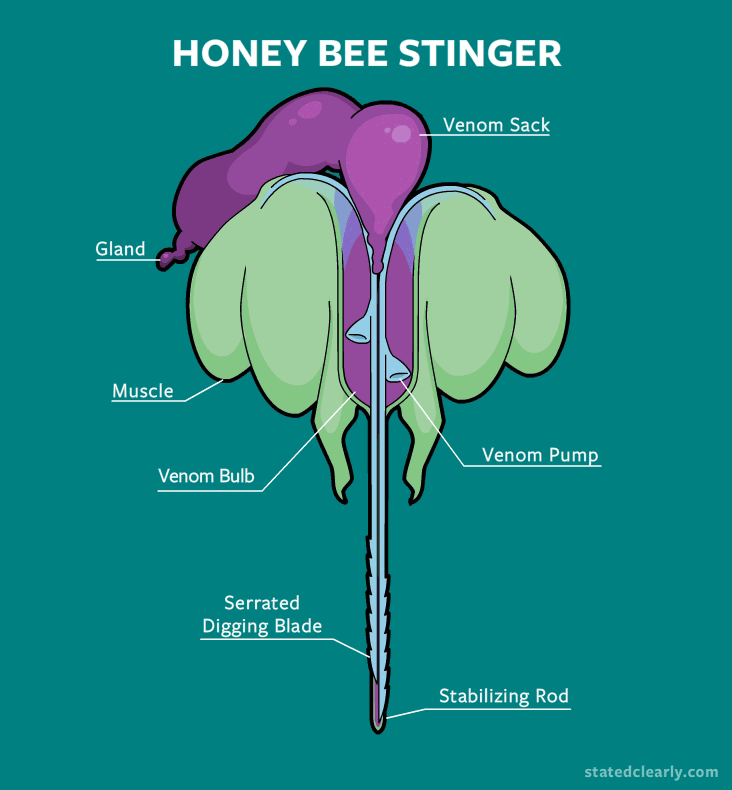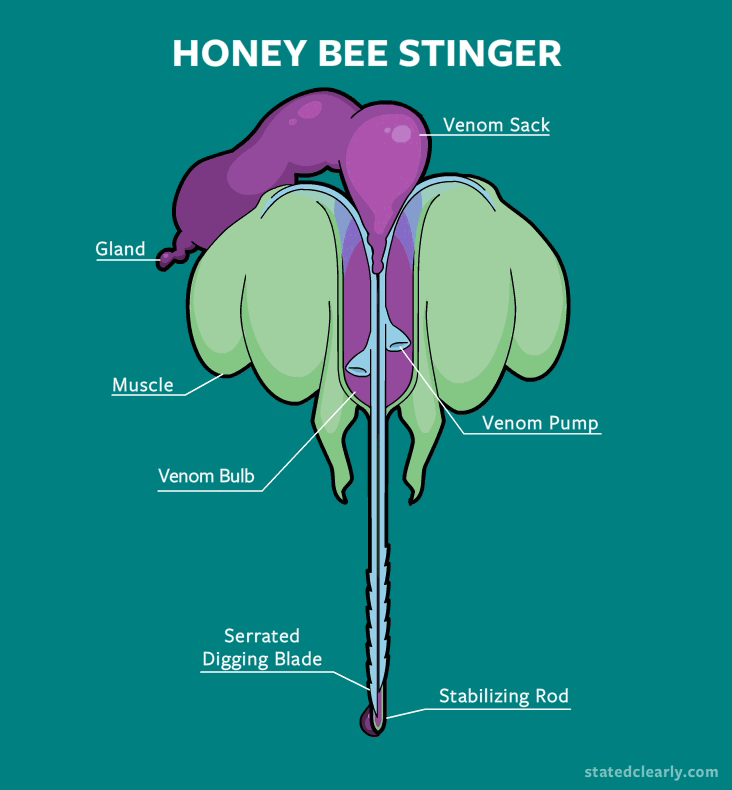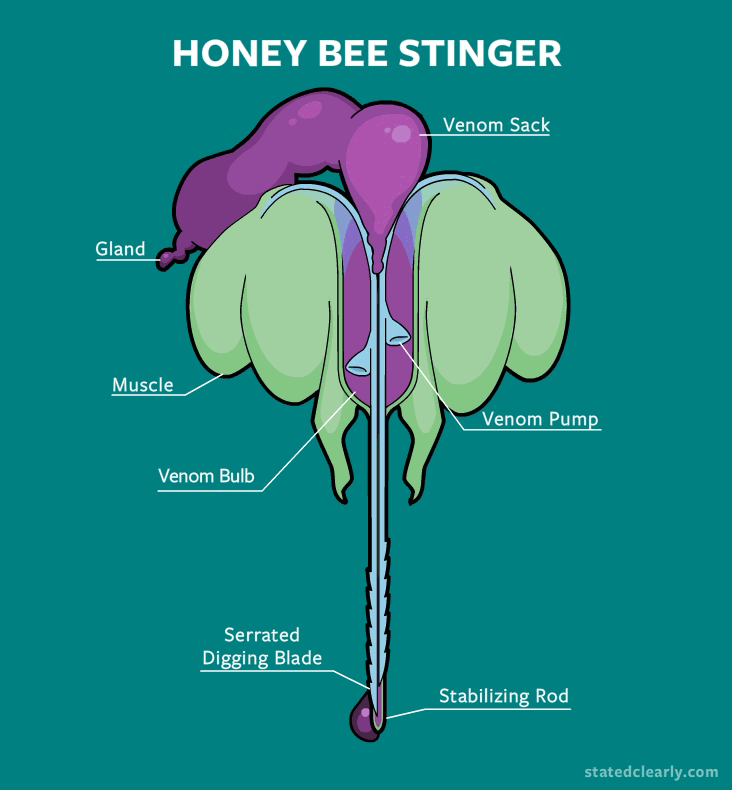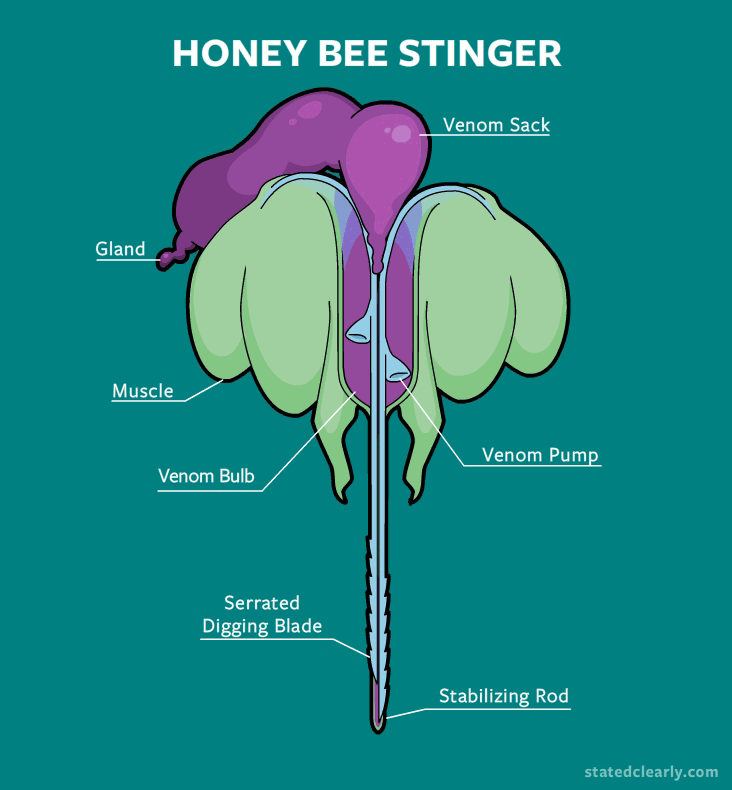
Rather than just sitting in your skin like a static fishhook, the stinger continues to pump venom into your skin long after the bee has flown away. This is made possible by the autonomous muscles and nerves in this pump, which work independently from the rest of the bee's nervous system. Up close in real life, this is pretty creepy and zombie-like.
If you don't pull the stinger out, the pump will keep on delivering every last drop of venom right into your skin. This mechanism is much more effective in terms of 'sting to volume of venom delivered' ratio compared to the short prick of a wasp.
Though this is actually an interesting comparison, because to us, a wasp's sting is just as painful as, if not even more painful than, a bee's. Wasps can also sting you several times without dying, multiplying the pain, so what gives?
Why does a bee have to die via this complicated, suicidal venom-pump mechanism whilst the wasps seem to do just fine with their non-barbed stingers?
Super-organisms, the immune system and colonial life
A simple answer to that would be: "bees die to defend the colony." Although compelling on the surface, this explanation would give a huge incentive for any individual bee to just hope that someone else sacrifices themselves other than them.
If you lived in a group of willingly sacrificial bees, it would be a very good strategy for you to be selfish and not sacrifice yourself. You can safely bet that someone else will take care of the cumbersome suicide, while you can kick back and enjoy the good life.
So why haven't selfish non-barbed-stinger bees become the norm?
To dig deeper, we need to look not just at the anatomy of the stinger itself but at the broader reproductive biology of the honey bee.
As I've mentioned previously on the blog, honey bee colonies have a distinctive social structure with the queen doing all the reproductive work, the female workers carrying out the honey production and the male drones providing the matching 16 chromosomes to fertilise bee eggs.
Drones don't do much for the colony besides that, and they don't have stingers like the workers or the queen.
The key point here is that the worker bees are reproductive dead ends for the wider colony (though there are some exceptions). So from an evolutionary perspective, it doesn't matter too much if they die, since they were never going to have kids anyway.
This does not mean that they are useless.
To see this, we can look at our own immune systems. For instance, immune cells inside your body known as neutrophils are just as suicidal as the worker bees. When bacteria invade your body, neutrophils migrate to the site of infection, unleash a whole cocktail of toxins onto the bacteria, and then proceed to die within 1-2 days.
In both cases, the suicidal nature of neutrophils and that of worker bees benefit the bigger entity that surrounds them. For worker bees, their death may save the colony from Winnie the Pooh, and the death of Neutrophils may save you from dying from a bacterial infection.
In both cases, the reproductive parts (the queen or the genitals) of each broader entity are preserved, whilst a disposable part (workers or neutrophils) which was never going to reproduce anyway, is destroyed. And for natural selection to do its thing, all we need are the reproductive parts to stay intact.
This line of thinking really does lead us to think of honey bee colonies as single super-organisms. Workers are just the cells of the organism that aren't going to have kids, so when they die, nothing too bad happens. Just like when you're skin cells or neutrophils die, which happens everyday by the way.
More importantly, with this distinction made, the "defence of the colony" explanation can be made to work if we reframe it slightly in terms of "defence of the queen" or "defence of the genitals."

This actually isn't the full story because as I'm sure you're aware, wasps also live in nests and so we could apply the same super-organism logic to them. Worker wasps ought to be just as dispensable as worker bees.
The reasons for why wasps are less suicidal than bees actually leads us down a different evolutionary path which we won't touch. But in short, wasps are just a more aggressive species than bees.

Ignoring this caveat, it still leaves the question of why super-organisms and colonial life evolved in the first place. I mean, we humans and nearly all other mammals certainly don't have a singular queen and a sterile caste of workers.
Group selection
Darwin himself recognised that the social insects (like wasps and bees) posed a problem for his theory of natural selection. Strict 'survival of the fittest' would seem to render eusociality, the extreme form of biological sociality found in these insects, impossible.
His own explanation for their existence involved an early form of group selection. We have since adapted it and modified it, though some biologists still get really triggered about group selection and deny its evolutionary importance. Ignoring that, here's how the theory would explain the evolution of eusociality.
Consider a group of non-social bees.
That is, a group that happens to live together but with no colonial social structure or any other social hierarchy that we see in honey bees today. So there is no queen and every bee can reproduce. Now imagine that one of these bees happens to be particularly nice and decides to share the food that it harvests with the rest of the group.
For the altruistic bee, this strategy is pretty terrible because she is giving away food that she has worked hard to collect. But for everyone else, this is great - they don't have to do as much work!
The receivers of free food can relax just a little and maybe spend their time doing other things. Perhaps even focussing on defending the group from predators with their new free time, instead of searching for food.
This bring us to the levels of selection in biology. On the individual level, it makes no sense to be altruistic; you're giving free food away for nothing in return. But on the group level, the presence of altruists is actually good as it can (in theory) increase the survivability of everyone, and therefore the reproductive fitness, of the entire group.
Comparatively, an all selfish group of bees may be more susceptible to predators as each individual's efforts are divided up between defence and searching for food. This group is less cohesive and easier to attack.
In this way, groups that have more altruists are more likely to survive, and the individuals within them are more likely to reproduce. The altruists among them in the group, pass on their altruistic traits, favouring the evolution of altruism and eusociality in groups. This is group selection.
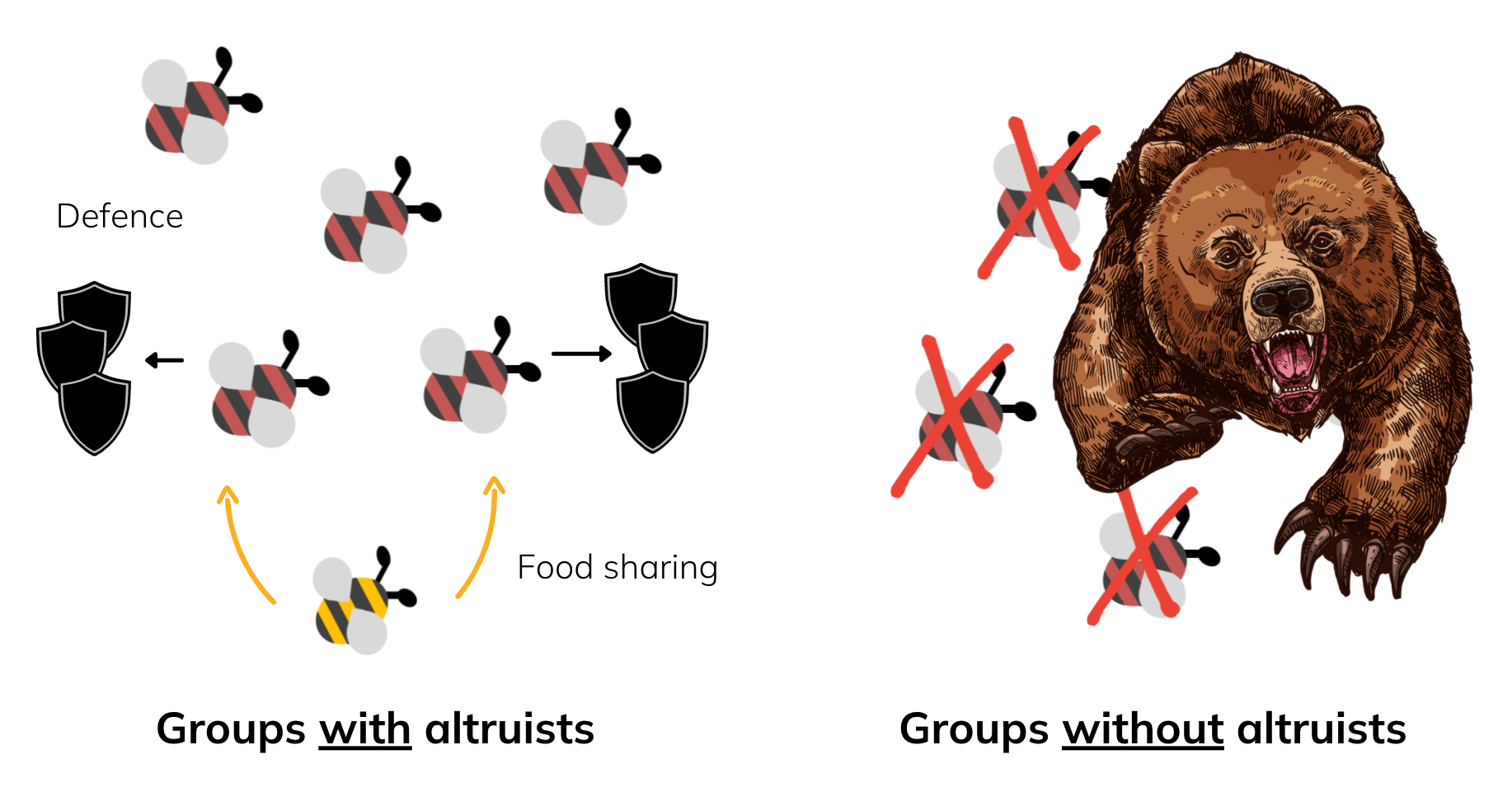
It is often naively said that altruistic traits can evolve because they favour the 'good of the group' or even more broadly the 'good of the species.' We need to be very careful with such phrases, especially the latter, because most individuals could not care less what happens to the rest of the species.
Modern biologists who support group selection are usually careful to point out the interactions between the levels of selection, like I did in my description above.
For this explanation to work, group level benefits (extra defence) have to outweigh individual level costs (giving away free food).
To highlight this nuance, group selection is commonly referred to as multi-level selection. We can also use maths to balance out the forces of individual-level selection and group-level selection. Hint: It's the Price equation again.
Group selection provides a possible mechanism for why altruistic traits, and therefore eusocial colonies like those of honey bees, may have evolved. But this doesn't explain why we only see eusociality in a select few species like wasps and bees, but not most others.
The nice story I told above about the evolution of altruism could just have easily been applied to humans. Yet we do not exist in eusocial colonies, so there must be something else going on.
Kin selection and biological altruism
In 1964, William D. Hamilton popularised a theory which became to be known as kin selection. In his groundbreaking paper, he provided a model to explain why eusociality was pretty much only seen in species of the order Hymenoptera which includes bees, wasps and ants.
Hamilton's first realisation was that Hymenopteran genetics is really weird. This was well known at the time, but he was the first person to truly understand the consequences of such a system.
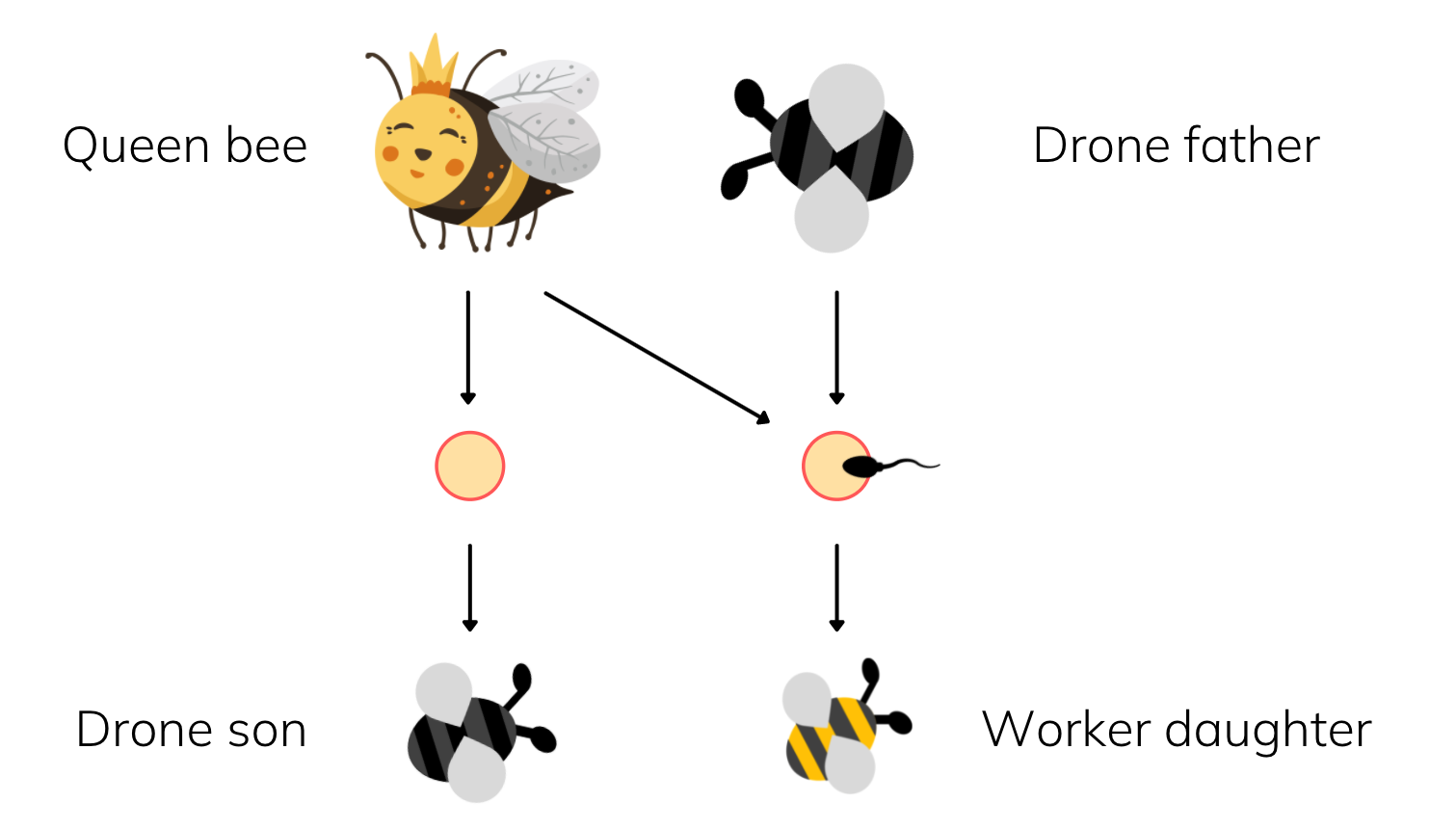
In Hymenoptera, all males come from unfertilised eggs, that is they only have a mother but no father. Females however, hatch out of fertilised eggs and get half their genes from each of their parents, much like us humans. This system of sex determination is known as haplodiploidy, as opposed to the diploid system found in mammals.
Now why does all this talk of genetics matter? Well, we can now assume that there is some gene that encodes for altruism.
When I was first learning about kin selection, this assumption really troubled me. Of course, there is no single gene that encodes for an altruistic trait. That's way too simple and completely ignores the role of cognition and the environment.
But what we can assume, which is considerably more reasonable, is that there is some gene that mildly inclines an individual to be more altruistic. With enough of these very slightly altruistic genes, the actual altruistic trait might emerge. For bees, we can imagine that one particular gene may make the barbs on its stinger ever so slightly longer.
Another confusion I had was the clash in the meaning of the word "altruism" in biology and in everyday conversation. Biological altruism refers to an individual giving up some of their reproductive fitness to help another individual reproduce.
These altruistic acts are typically not done consciously, they have simple been formed by natural selection to act in this way. For instance, your neutrophils do not consciously commit suicide, they have just been shaped to do that by evolution. And of course you don't go out thinking "Hm yes I will save you two from drowning because you are each 5/8 related to me." Although maybe one biologist did actually think like this.
The best analogy I can think of is the following. To catch a ball in the air requires your brain to do some pretty complex calculus, but you never once think of all this consciously. Same with all this relatedness business.
With that cleared up, Hamilton's insight was now two-fold:
- You can spread your genes 'through' your relatives.
- The haplodiploid system skews the relatedness between sisters.
Let's go through each of these individually.
First, we need to take the 'gene's eye view' of evolution for all of this to make sense. So imagine that you are an altruistic gene and that your goal is to maximise your own personal fitness. Irrespective of your downstream function, you only have your own selfish interests at heart. That is, you only care about creating more copies of yourself.
This perspective was precisely why Richard Dawkins named his book The Selfish Gene. And although there is a lot of anthropomorphising to make the analogy work, it is useful to understand how kin selection operates.
Here's where the apparent altruism comes in.
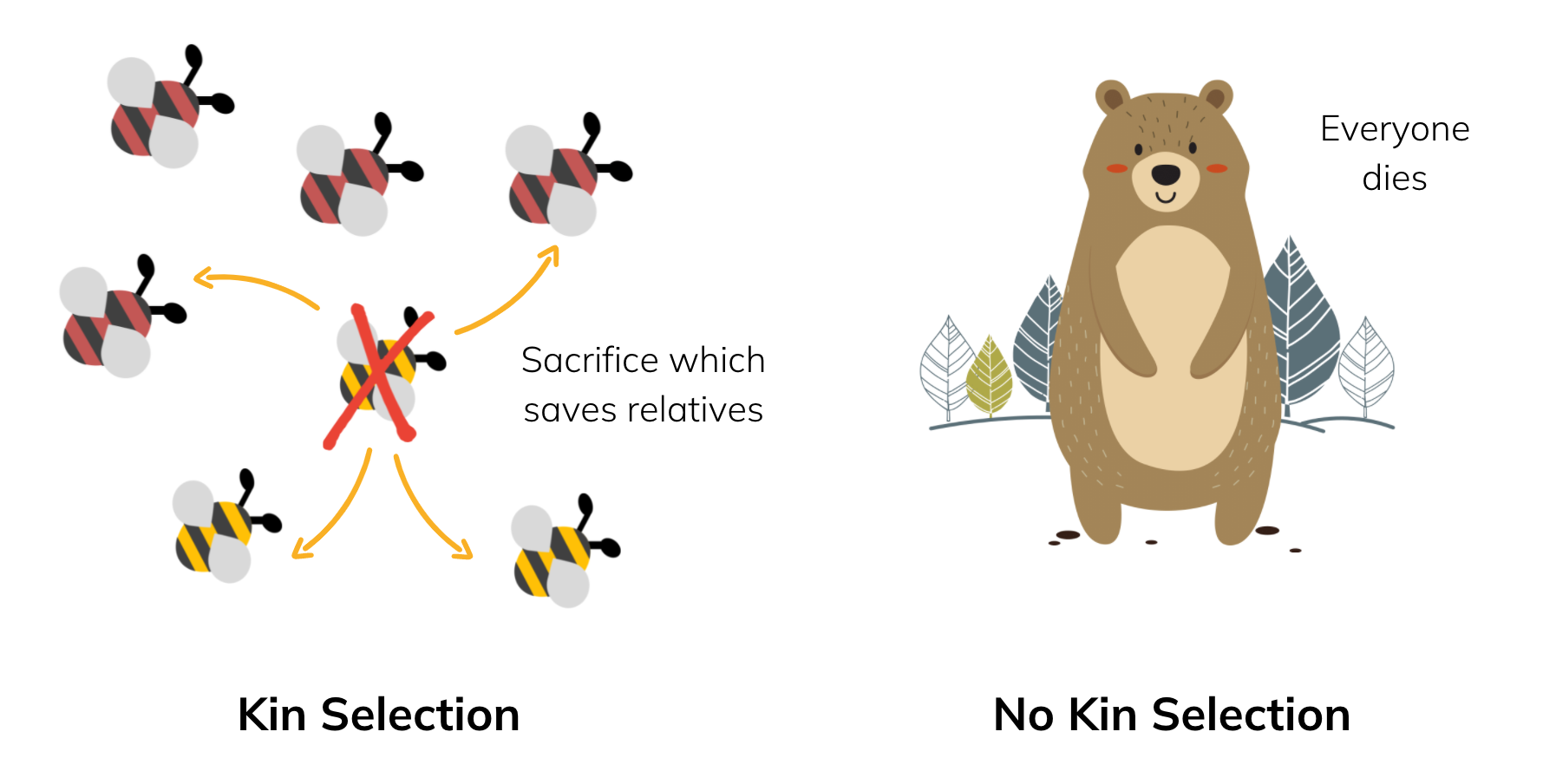
Imagine that there is some external pressure, say a predator, that will cause everyone in your group to die if nothing is done. If however, one of you sacrifices yourself to the scary predator, you can save everyone else.
As a slightly-altruistic gene, you are more inclined to make the individual that you live inside conduct this sacrificial act. From the group selection perspective, this action never made any sense for the individual. But from the gene's eye view, this can actually be beneficial if you happen to save some other individuals that also contain a copy of yourself.
This is a way of replicating yourself through others instead of directly.
The individuals you save which do not contain a copy of you are essentially wasted energy; you're saving them for no reason. So for kin selection to work, it's very important that the individuals that you do happen to save are closely related to you, in order to maximise your chances of saving copies of yourself. This is the 'kin' part in kin selection.
Notice that when this reproduction through relatives occurs, the gene for altruism spreads and the descendant individuals will end up slightly more altruistic.
Indirect fitness and the haplodiploidy hypothesis
The easiest way to conceptualise this process is through the concept of indirect fitness. This is a formal way of expressing "passing on your genes through relatives" as opposed to having kids yourself. Reflecting upon this fact, another 20th century biologist, J.B.S. Haldane famously said:
I would gladly give up my life for two brothers or eight cousins.
This is because in humans, you share half of your genes with your siblings (and each of your parents) and an eighth of your genes with first-cousins. In Hymenoptera however, the situation is more complex precisely because of their strange genetics.
Let's have another thought experiment to see this in action. Imagine that you are now a female bee and that you have two options during mating season:
- Help your mum to have more babies (producing more siblings in relation to you).
- Have your own kids.
And rather than jumping to option 2 which intuitively sounds more reasonable, we can use our new tools of kin selection and indirect fitness to work out which one is actually better for you. Remember that we want to maximise the genes that we pass on, so higher relatedness to new individuals produced is better.
If you have a child as a Hymenopteran female, no matter whether it turns out to be male or female, you will have passed on half of your genetic material. Nothing too weird yet, this is the same as us.
If on the other hand, you help your mum to have another daughter, how related are you to your new sister?
Well, both of you have the same father who has passed on his entire set of genes to both of you. And you each have gotten some random half of your mother's genes (via meiosis). So adding all of that up, you actually share three-quarters of your genes with your sister not half!
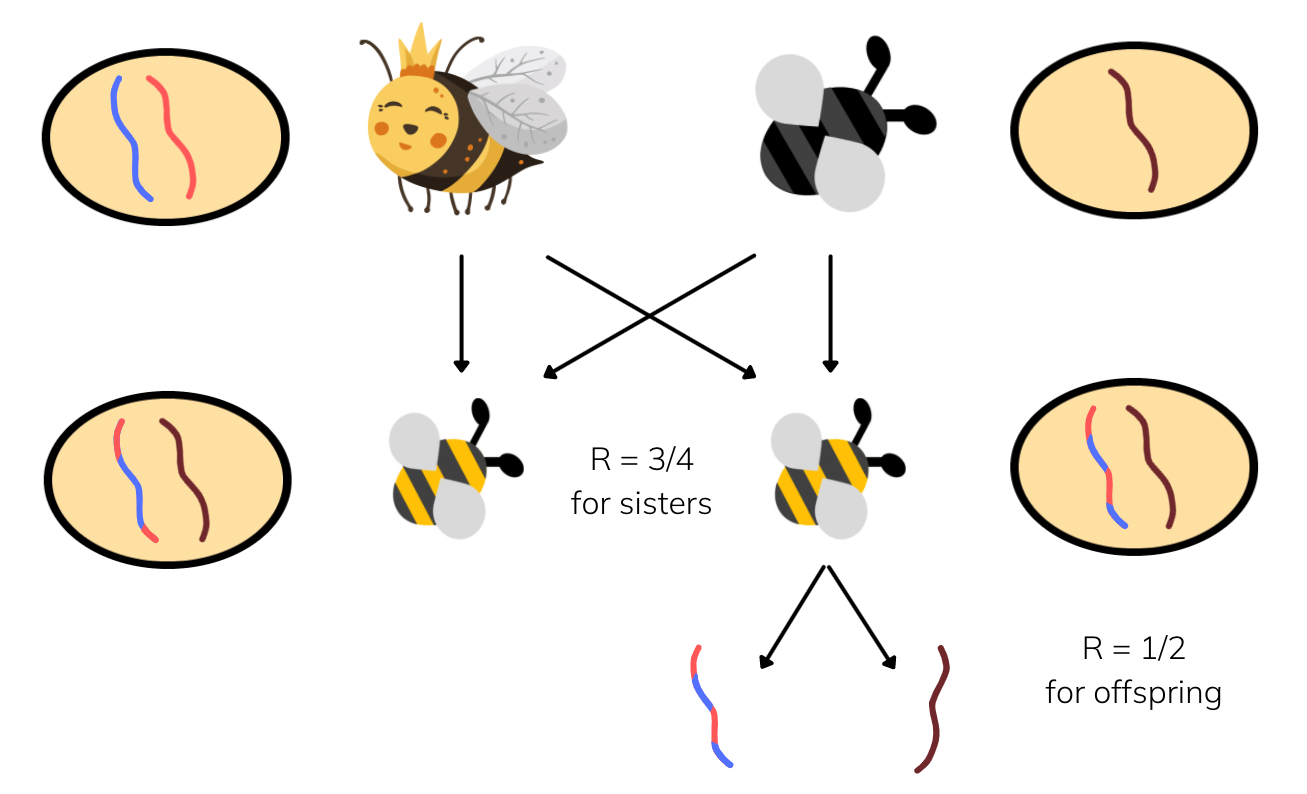
For those of you familiar with genetics: Male Hymenopterans do not undergo any meiosis, so their gametes are all essentially clones of themselves. Females do undergo standard meiosis which gives them the standard R=1/2 relatedness to all offspring. Combining these together gives R=3/4 between sisters, assuming a common father.
With such high relatedness between sisters (3/4 > 1/2), it actually makes more sense to use your mother as a 'sister-producing factory' to maximise the genes that you pass on as a female bee.
This is the indirect fitness concept at its finest and it also gives a very good explanation for why worker bees are so disposable. All that matters to the workers is that this sister-producing factory (the queen) is preserved. They don't care about reproducing on their own.
Thus, the 3/4 relatedness between Hymenopteran sisters gives the additional boost required to favour the evolution of eusociality via kin selection.
This is Hamilton's haplodiploidy hypothesis, and it is a pretty compelling explanation for why eusociality seems to have mostly cropped up in only one order of life, one which happens to have haplodiploid genetics.
The haplodiploidy hypothesis isn't perfect
The haplodiploidy hypothesis, though nice, is not without its problems. Firstly, there are plenty of species with haplodiploid genetics which do not form eusocial colonies. This includes many species of solitary bees, wasps and some rotifers. So being a haplodiploid species is not synonymous with eusociality.
Secondly, there are also instances of diploid species that are social. One is the very strange naked mole rat which I think definitely earns the title of "strangest mammal alive today".
Thirdly, the haplodiploidy hypothesis only works if all sisters share the same father and if a queen is biased to produce more daughters than sons. Both of these conditions are not often satisfied in modern honey bee species. Queens often mate with more than one male (resulting in multiple fathers) and sex ratios are usually equal in most species, not female-biased.
Many additional models have been proposed to fix these problems, and it is an active area of research today. One simple fix is having some colonies with female-biased sex ratios and others with male-biased sex ratios. Overall, this keeps a 50/50 balance in the whole population but allows for kin selection to operate locally with the 3/4 relatedness boost within certain colonies.
Despite its problems, the core idea of Hymenopteran genetics favouring the evolution of eusociality seems to hold up pretty well.
Conclusion
So why do bees die when they sting you? Well, it ultimately depends on which biologist you ask.
Perhaps because they're disposable parts of a larger super-organism which has evolved by multi-level selection. Perhaps because they're happy to die for their sister-producing factory known as the queen, fuelled by kin selection and the haplodiploid genetics of the Hymenoptera.
Nothing is certain because we don't have a time machine to trace the evolution of bees. Biologists are certainly still debating about it and neither kin nor group selection has reached unanimous consensus.
Jake

Member discussion: